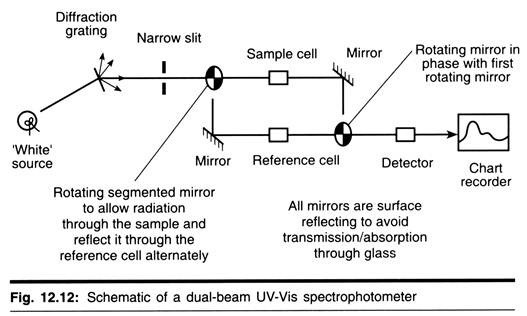
ADVERTISEMENTS:
V tomto článku se budeme zabývat buněčnými strukturami řas pomocí diagramů.
I. Buněčná stěna:
Buněčná stěna většiny řas je celulózní. Obsahuje také hemicelulózu, sliz, pektin a další látky, jako je kyselina alginová, fukoidin, fucin, uhličitan vápenatý, oxid křemičitý atd. v různých kombinacích u různých skupin řas. Elektronmikroskopické studie ukazují, že celulózová stěna je složena z celulózových mikrofibril různé tloušťky, které zůstávají různě orientované v zrnité matrici.
Buněčná stěna diatomů je silicifikovaná a vykazuje charakteristické sekundární struktury. U Cyanophyceae je buněčná stěna tvořena mukopeptidem sestávajícím převážně z peptidu několika aminokyselin kovalentně vázaných na aminocukry, glukosamin a kyselinu muramovou.
DOPLŇKY:
Tento mukopeptidový komplex je také přítomen v buněčné stěně bakterií. Pravá buněčná stěna chybí u některých řas, jako jsou Gymnodinium a Pyramimonas. Místo toho mají hraniční membránu známou jako pelikulu.
II. Protoplast:
Protoplazmatický obsah buňky se nazývá protoplast. Eukaryotický řasový protoplast je obklopen lipoproteinovou vnější hranicí, nazývanou buněčná membrána, a skládá se z jednoho nebo více obvykle kulovitých nebo elipsoidních jader a cytoplazmy.
Buněčná membrána se skládá z lipidů a bílkovin a má podobně jako všechny ostatní biologické membránové systémy tekutou mozaiku. Je velmi tenká a pružná a selektivně propustná. Řídí průchod látek dovnitř a ven z buněk.
Jádro:
U všech eukaryotických řas (Chlorophyceae aj.) je jádro dobře organizované kulovité nebo eliptické těleso. Zůstává obklopeno výraznou jadernou membránou. Vnitřní stranu zaujímá chromatinové retikulum uložené v matrici zvané karyolymfa. Jaderná membrána je dvouvrstvá. Vnější membrána je souvislá s endoplazmatickým retikulem.
Každé jádro obsahuje jedno nebo více nukleol nebo endosomů. Počet nukleolů se u různých řas liší. Může být jedno, dvě nebo více. Chromozomy mohou mít lokalizovanou nebo rozptýlenou centromeru. Počet chromozomů se u jednotlivých druhů liší – nejnižší počet je n=2 (např. Porphyra linearis), nejvyšší n=592 (Netrium digitali).
V eukaryotické buňce řas se nacházejí buněčné organely vázané na membránu, jako jsou chloroplasty, mitochondrie, Golgiho aparát, endoplazmatické retikulum a v některých případech oční skvrna nebo stigma.
U prokaryotické řasové buňky (příslušníci sinic) není jádro ohraničeno žádnou membránou. Místo toho je protoplast diferencován na vnější periferní chromoplazmu obsahující fotosyntetické pigmenty a vnitřní bezbarvou centroplazmu, kde se genetický materiál nenachází v membránou vázaném jádře a vlákna DNA se nespojují s histony za vzniku chromozomů.
Centroplazma tedy u Cyanophyceae představuje počáteční jádro. Jádro u Dinophyceae také není skutečně eukaryotické, je sice vázané na membránu, ale chromozomy a mitotický aparát chybí. Cytoplazma řasové buňky se dělí na buněčné organely a cytosol.
1. Chloroplast:
Chloroplasty jsou velmi výrazným znakem buněk řas. Jsou nositeli fotosyntetických pigmentů. Jedná se o dvoumembránovou strukturu.
U různých druhů řas jsou známy různé formy chloroplastů, z nichž se obvykle rozeznává osm hlavních typů : pohárkovité (např. Chlamydomonas a Volvox), diskovité (např. Chara, Vaucheria a centrické diatomy), parietální (např, Chaetophorales, Phaeophyceae, Rhodo- phyceae, mnohé Chrysophyceae a zpeřené diatomy), pásovité nebo C-tvarované (např. Ulothrix), spirálovité (např. Spirogyra), síťovité (např. Oedogonium, Hydrodictyon a Cladophora), hvězdicovité (např. Zygnema) a žebernaté (např. Volvocales). Základní struktura chloroplastu je téměř podobná v celé rostlinné říši.
PŘÍKLADY:
V chloroplastu existují tři hlavní strukturní oblasti:
1. Obal tvořený dvěma membránami s uzavřeným prostorem.
2. Pohyblivá stroma obsahující rozpustné enzymy pro metabolismus, syntézu bílkovin a skladování škrobu a
3. Vysoce organizované vnitřní lamelární membrány obsahující pigmenty a podílející se na zachycování a přenosu energie.
PŘÍČINY:
Vnitřní lamelární systém tvoří disky, které jsou poskládány na sebe jako hromádky mincí a tvoří grana. Každý disk je váček nebo vezikula a označuje se jako thylakoid. Každý tylakoid uzavírá intertylakoidní prostor.
Tylakoidní systém tvoří jedinou složitou dutinu, která je od stromatu oddělena tylakoidní membránou. V tylakoidních membránách jsou zabudovány pigmentové systémy a přenašeče elektronů, které provádějí světelnou fázi fotosyntézy.
U Cyanophyceae nejsou tylakoidy uzavřeny ve skupinách vázaných na membránu a tvoří chloroplasty, místo toho leží volně v cytoplazmě. Tylakoidy jsou místem výskytu chlorofylu a a na jejich povrchu se vyskytují také akcesorní pigmenty v podobě malých vezikul zvaných fykobilisomy.
a. Pigmentace:
PIRIGMENTY:
Pigmenty, které zajišťují vlastní barvu thallusu, jsou různého typu:
i. Chlorofyly:
V řasách se vyskytuje pět typů chlorofylů: Chi a, b, c, d a e. Z nich je chlorofyl a přítomen ve všech skupinách řas. Chlorofyl b se vyskytuje pouze u Chlorophyceae, chlorofyl c u Phaeophyceae, Cryptophyceae, Bacillariophyceae a Chrysophyceae, chlorofyl d u některých červených řas a chlorofyl e u některých Xanthophyceae.
ii. Karoteinoidy:
PŘEDPOKLADY:
Karoten a xantofyl společně tvoří karotenoidy. Jsou to akcesorické fotosyntetické pigmenty.
V řasách se vyskytuje pět typů karotenů: α-karoten v Chlorophyceae, Cryptophyceae a Rhodophyceae; β-karoten ve všech skupinách řas s výjimkou Cryptophyceae; c-karoten v Chlorophyceae; e-karoten v Bacillariophyceae, Cryptophyceae, Phaeophyceae a Cyanophyceae a flavacen v. Cyanophyceae.
iii. Xantofyly:
V řasách se vyskytuje několik typů xantofylů. Z nich lutein, violaxanthin a neoxanthin se vyskytují u příslušníků Chlorophyceae a Phaeophyceae. Fukoxanthin je hlavním xantofylovým pigmentem u Phaeophyceae a Bacillariophyceae, zatímco myxoxanthophyll, myxoxanthin a oscilloxan- thin se vyskytují pouze u Cyanophyceae.
iv. Fykobiliny:
Fykobiliny jsou ve vodě rozpustné lineární tetr’apyroly. Jsou to biliproteiny červené (fykoerytrin) nebo modré (fykocyanin) barvy. Vyskytují se pouze u Rhodophyceae a Cyanophyceae. Fungují jako akcesorické pigmenty tím, že absorbují a přenášejí světelnou energii do reakčního centra.
Doporučení:
b. Pyrenoidy:
Pyrenoidy jsou bílkovinná tělíska přítomná v chloroplastech nebo chromatoforech – právě to je charakteristické pro chloroplasty řas. Obvykle jsou spojeny se syntézou a skladováním škrobu. U Bacillariophyceae se v nich hromadí lipidy. Počet pyrenoidů může být jeden (např. Chlamydomonas) nebo více než jeden (např. Oedogonium) na jeden chromatofor.
2. Mitochondrie:
Mitochondrie se nacházejí ve všech buňkách řas kromě Cyanophyceae. Každá mitochondrie je obklopena dvojitým membránovým obalem. Vnitřní membrána rostlinných mitochondrií uzavírá vodní matrici rozpuštěných látek, rozpustných enzymů a mitochondriální glukózy.
Vnitřní membrána je větší než vnější membrána a podléhá invaginaci, čímž vznikají váčkovité krysty různého tvaru a počtu – obvykle s úzkým hrdlem. Celá mitochondrie je opět obklopena vnější membránou ležící v těsné blízkosti vnitřní a ponechává mezimembránový prostor, který je spojitý s mezikristálním prostorem.
Matrix je jemně zrnitá a vysoce proteinová. Organella má semiautonomní povahu, protože obsahuje vlastní kruhovou DNA a ribozomy, s jejichž pomocí může syntetizovat některé své bílkoviny. V každé buňce je obvykle více než jedna mitochondrie, ale u Micromonas (Chlorophyceae) obsahuje každá buňka jedinou mitochondrii
ADVERTISEMENTY:
Buňky sinic mitochondrie postrádají. Cytoplazmatická membrána je místem biochemických funkcí, které jsou u eukaryotických buněk obvykle spojeny s přesně vymezenými membránovými organelami. Podobně jako u bakterií se buněčná membrána invaginuje a vytváří strukturu zvanou mezo-som, kde jsou lokalizovány dýchací enzymy.
3. Endoplazmatické retikulum (ER):
Elektronově mikroskopické studie ukazují, že buňky řas obsahují rozsáhlou membránovou síť vzájemně propojených kanálků a cisteren (zploštělých váčků), která se nazývá endoplazmatické retikulum. Membrány ER procházejí celou cytoplazmou.
Retikulum se skládá ze vzájemně propojených paralelních cisteren spojených s ribozomy, které jsou připojeny k cytoplazmatické straně membrány. Tato forma ER je známá jako hrubé endoplazmatické retikulum (RER), které je hlavním místem syntézy proteinů. Naopak membrány ER, které nenesou ribozom, se nazývají hladké endoplazmatické retikulum (SER).
4. Diktyozomy neboli Golgiho aparát:
Diktyozomy neboli Golgiho tělíska se nacházejí ve všech buňkách řas s výjimkou sinic a lze je pozorovat pod elektronovým mikroskopem. Golgiho aparát je součástí endomembránového systému buňky a zřejmě slouží jako mezičlánek mezi endoplazmatickým retikulem a plazmatickou membránou.
Golgiho tělíska se mohou nacházet v oblasti jádra (např. Chlamydomonas), v blízkosti plastidů (např. diatom a Bulbochaete) nebo se mohou nacházet kdekoli v buňce. Golgiho tělíska se skládají ze 2-20 plochých vezikul, které jsou uspořádány do hromádek.
Každá hromádka se nazývá dictyosom. Všechny diktyozomy dohromady tvoří Golgiho aparát. Funguje při balení materiálů určených k exportu do vnější části buňky. Je také zodpovědný za tvorbu nové plazmatické membrány, která podporuje růst nebo nahrazuje ztracenou.
5. Oční skvrna nebo stigma:
Pohyblivé vegetativní a reprodukční buňky řas mají v přední, střední nebo zadní části buňky pigmentovou skvrnu, známou jako oční skvrna nebo stigma (obr. 3.12). Podílí se přímo nebo nepřímo na vnímání světla. Stigma se obvykle nachází uvnitř tylakoidů probíhajících podélně přes oční skvrnu mezi dvěma řadami granulí.

6. Vakuoly:
Téměř všechny buňky řas, s výjimkou příslušníků Cyanophyceae, mají jednu nebo více vakuol. Každá vakuola je ohraničena samostatnou membránou zvanou tonoplast.
U pohyblivých forem se vyskytují tři typy vakuol:
(i) Jednoduché vakuoly:
Jsou velmi malé a vykazují periodické smršťování a rozpínání. Nazývají se také kontraktilní vakuoly. Vyhazují metabolické odpady buněk. Regulují také obsah vody v buňce tím, že v krátkém intervalu vypouštějí její přebytečné množství. Mají tedy sekreční funkci.
(ii) Složené vakuoly:
Tato vlastnost je charakteristická pro Dinophyceae a Euglenophyceae. Skládá se z trubicovitého cytofaryngu, velkého rezervoáru a skupiny vakuol různé velikosti. Vakuoly plní uvnitř buňky funkci osmoregulace. Někdy se ve vakuolách ukládají také rezervní potravní látky, jako je laminarin a chrysolaminarin.
(iii) plynové vakuoly:
V buňkách příslušníků Cyanophyceae jsou dutiny obsahující plyn, které se vyskytují jako hromádky malých průhledných válců stejného průměru. Jejich stěny jsou volně propustné pro plyny. Plynové vakuoly dodávají planktonním formám vztlak a slouží také jako ochranné clony proti dopadajícímu jasnému světlu.
7. Bičíky:
Motilující vegetativní nebo reprodukční buňky jsou přítomny u všech skupin řas kromě Cyanophyceae a Rtiodophyceae. Pohybu je dosaženo tlukotem malých vláknitých nebo nitkovitých protoplazmatických přívěsků, které se nazývají bičíky. Liší se počtem, délkou, polohou a přítomností nebo nepřítomností chloupků v různém počtu. Jejich počet se pohybuje od jednoho do čtyř nebo mnoha.
Jsou převážně dvojího typu:
1. Bičíkaté nebo akronematické:
Jsou to bezvlasé bičíky s hladkým povrchem (obr. 3.13A, obr. 3.14A) a
2. Bičíkaté neboli pleuronematické:
Ty mají jednu nebo více řad postranních jemných vláknitých chloupků známých jako mastigonemy nebo flimery
Dále se dělí na:
a. Pantonematické. Kde jsou mastigonemy uspořádány ve dvou protilehlých řadách (obr. 3.13B).
b. Pantoakronematické. Pantonematický bičík s terminálním vláknem se nazývá pantoakronematický (obr. 3.13C).
c. Stichonematické. Má jednostranné mastigonemy (obr. 3.13D).

Pokud je počet bičíků na buňku větší než jeden a shodný, označuje se jako izokont, pokud je rozdílný, označuje se jako heterokont.
Bičíkaté kořeny u řas:
Bičíky jsou extrémně jemné, hyalinní výrůstky cytoplazmy. Běžně se na bázi každého bičíku nachází jediná granule (obr. 3.15A), blefaroplast nebo bazální tělísko. Pokud má řasová buňka pevnou stěnu, bičík vystupuje pórem.
Každý bičík má centrální nebo axiální tenké vlákno, axoném. Axoném je obklopen cytoplazmatickou membránou nebo pochvou tvořenou prodloužením buněčné nebo plazmatické membrány. Plášť končí těsně před vrcholem bičíku. Vrcholová obnažená část axonemu se nazývá koncová část.
Na příčném řezu bičíkem (obr. 3.15B) jsou vidět dvě centrální singletová vlákna obklopená devíti periferními dubletovými vlákny. Každá fibrila je pokryta membránou a dvě centrální jsou dále pokryty další membránou.

