Morbillivirukset ovat erittäin tarttuvia taudinaiheuttajia, ja ne aiheuttavat erilaisia taudinpurkauksia altistumattomissa väestöissä (Pfeffermann et al., 2018). Ne kuuluvat järjestykseen Mononegavirales ja perheeseen Paramyxoviridae, ja niille on ominaista segmentoimaton, lineaarinen, negatiivisjuosteinen RNA-genomi (Lamb ja Parks, 2013). Morbillivirukset erottuvat siitä, että ne aiheuttavat keskivaikeita tai vaikeita hengitystie-, ruoansulatuskanava-, immunosuppressio- ja/tai neurologisia sairauksia monenlaisissa isännissä, kuten ihmisissä (tuhkarokkovirus), lihansyöjissä (koiran morbillivirus, aiemmin canine distemper-virus), nautaeläimissä (rinderpest-virus), delfiineissä ja pyöriäisissä sekä muissa luonnonvaraisissa eläimissä elävissä eläinlajeissa (Lamb and Parks, 2013; Martinez-Gutierrez and Ruiz-Saenz, 2016).
Tuhkarokkovirusta (MeV) ja koiran morbillivirusta (CDV) pidetään tämän perheen tarttuvimpina viruksina (De Vries et al., 2015), ja CDV:n suuren tarttumispotentiaalin sekä sen lajien välisen tarttumispotentiaalin vuoksi maailmanlaajuiset terveys- ja luonnonsuojeluviranomaiset ovat erittäin huolissaan CDV:n roolista uhanalaisten lajien suojelussa ja mahdollisesta ”hypystä” eläimistä ihmisiin (Terio ja Craft, 2013; Ohishi et al., 2014). Kotieläiminä pidetyt koirat ovat CDV:n pääasiallinen isäntä, ja niitä voitaisiin pitää myös muiden nisäkkäiden reservoirina (Suzuki ym., 2015; Duque-valencia ym., 2019); CDV:n biologian perusteella myös ihmisistä voisi kuitenkin tulla potentiaalinen kohde (Cosby ja Weir, 2018; Rendon-Marin ym, 2019).
Yritettäessä ymmärtää CDV:n mahdollista riskiä tarttua ihmisiin on tarpeen kerätä kaikki olemassa oleva todistusaineisto; ja tutkimus tämän taudinaiheuttajan alkuperästä ja leviämisestä koirapopulaatiossa voisi olla tärkeä avain tämän prosessin ymmärtämiseen. Hiljattain International Journal of Paleopathology -lehdessä julkaistu artikkeli herätti keskustelua CDV:n evolutiivisesta alkuperästä. Siinä päädytään siihen, että CDV sai alkunsa pandemisena taudinaiheuttajana Etelä-Amerikassa MeV:n tartuttua ja sopeuduttua koiriin Etelä-Amerikan kolonisaatiokaudella. Tämä tulos saatiin tieteidenvälisellä lähestymistavalla, joka omaksuttiin syntetisoimalla paleopatologinen analyysi 96 prekolumbiaanisesta koirasta (750-1470 CE), jotka olivat peräisin Weyanoken vanhasta kaupungista Virginiassa, historiallisista raporteista, molekyylianalyysistä ja morbillivirusepidemiologiasta (Uhl et al., 2019).
Amerikasta peräisin olevat alkuperäiset koirapopulaatiot lähes katosivat kolonisaatioajan jälkeen, ja mantereelle tuotiin eurooppalaisia ja euraasialaisia koiria, jolloin sen amerikkalaisista edeltäjistä jäi vain vähän geneettistä taustaa (Ni Leathlobhair ym., 2018). Toinen tärkeä tekijä, joka on syytä ottaa huomioon, on se, että myös ”tuntemattomia” tauteja on voitu tuoda maahan, mikä vaikeuttaa uusien taudinaiheuttajien alkuperän jäljittämistä. Lisäksi kotikoirien ja jopa ihmispopulaatioiden keinotekoinen valintapaine erityisesti kolonisaation aikana on voinut lisätä tautien esiintyvyyttä ja siten rajoittaa geneettistä vaihtelua (Ostrander et al., 2017), mikä puolestaan saattoi merkitä vähemmän tehokasta vastetta taudinaiheuttajia vastaan.
Muista näistä ”uusista” taudinaiheuttajista/sairauksista CDV:n kuvasi ensimmäisen kerran Antonio de Ulloa y de la Torre-Giral vuonna 1746 koirien sairastamana tautina Quiton alueella ja muualla Etelä-Amerikassa, ja siitä raportoitiin pian sen jälkeen myös Euroopassa. CDV todettiin Espanjassa vuonna 1760, jolloin Madridissa sattui 900 kuolemantapausta yhden päivän aikana, ja kolme vuotta myöhemmin, eli vuosina 1764 ja 1770, se oli saavuttanut Ison-Britannian ja Italian (Blancou, 2004). Edward Jenner raportoi myöhemmin 1800-luvun alussa viruksen tarttuvuudesta ja pentujen suuremmasta alttiudesta aikuisiin koiriin verrattuna. Hän vertasi niiden tarttuvuutta MeV:n tarttuvuuteen ja havaitsi, että eloonjääneet olivat suojassa myöhemmiltä tartunnoilta (Jenner, 1809; Nambulli et al., 2016).
Lyhyesti sanottuna eurooppalaisten pioneerien saavuttua 1400-luvulla uusista tartuntataudeista tuli luultavasti kolonisaation tuhoisin seuraus, koska Amerikan alkuperäisväestöllä ei ollut aiempaa altistumista patogeeneille, jotka olivat yleistyneet Euroopassa (Walker et al., 2015). Useat tuhkarokkoepidemiat tuhosivatkin Amerikan alkuperäisväestöjä (Walker et al., 2015; Nambulli et al., 2016). Uhl et al. raportoivat paleopatologisten, historiallisten, molekulaaristen ja epidemiologisten todisteiden yhdistetyn lähestymistavan kautta, että Amerikan alkuperäisväestöissä esiintyneet vakavat MeV-epidemiat edesauttoivat MeV:n hyppyä Etelä-Amerikan kaupunkiympäristöjen suuriin kotikoirapopulaatioihin ja viruksen sopeutumista endeemiseksi CDV:ksi (Uhl et al., 2019). Historialliset tiedot voivat myös todistaa, että muutama vuosi tuon Etelä-Amerikan koiriin sopeutumisen jälkeen CDV kulkeutui Eurooppaan vuonna 1760, jossa se aiheutti aluksi laajoja epidemioita, joihin liittyi korkea kuolleisuus, ennen kuin siitä tuli endeeminen (Jenner, 1809).
Evoluutioennusteisiin liittyvä molekulaarinen fylogeografia ja aika viimeisimpään yhteiseen esi-isään (tMRCA) laskettiin kuitenkin CDV:n alkuperälle Yhdysvalloissa 1880-luvulla (95 %:n korkein posteriorinen tiheys, 1858-1913) (Panzera ym, 2015), mikä on selvästi ristiriidassa viruksen kuvauksen kanssa Euroopassa 1700-luvulla. Tähän hypoteesiin johtaneita sekvenssianalyysejä on tutkittava huolellisesti, koska tässä molekulaarisen fylogeografian rekonstruktiossa käytetyt sekvenssit ovat puolueellisia ja niiden saatavuus on rajallinen. Lisäksi monet alkuperäiset esivanhempien sekvenssit ovat kadonneet CDV:n ja muiden morbillivirusten viruksen RNA-genomin hitauden vuoksi. Nämä tekijät ovat antaneet aihetta kyseenalaistaa nykyisten tMRCA-laskelmien käyttökelpoisuuden RNA-viruksille (Sharp ja Simmonds, 2011; Nambulli ym., 2016).
Uhl ym. mukaan, morbillivirus on voinut saada alkunsa karjasta noin 376 eaa. ”vanhalla mantereella” (kuva 1), ja eläinten kesyttämisellä on saattanut olla merkittävä vaikutus lajien välisiin tapahtumiin, mikä todennäköisesti jäljittää MeV:n syntymisen alkamisajankohdan noin vuoteen 900 eaa. (Uhl et al., 2019). Toisin kuin nykyiset CDV:n fylogeneettiset rekonstruktiot, MeV:n divergenssi saa vahvaa tukea rennon kellon Bayesin fylogeneettisestä analyysistä. MeV:n ja karjaruttoviruksen välisen eroamisajankohdan oli osoitettu tapahtuneen noin yhdellätoista kahdellatoista vuosisadalla (Furuse ym., 2010). Muut molekulaariset tiedot, kuten Brasiliasta peräisin olevissa lepakoissa kiertävän uuden morbilliviruksen (joka on läheistä sukua CDV:lle ja PDV:lle) esiintyminen (DrMV), antavat mahdollisuuden spekuloida, että CDV:llä ja DrMV:llä saattaisi olla yhteinen eteläamerikkalainen esi-isä (Drexler ym., 2012), mikä epäsuorasti tukee ajatusta CDV:n varhaisesta eteläamerikkalaisesta alkuperästä.
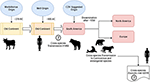
KUVA1. Kaavamainen esitys mahdollisesta koiran morbilliviruksen (CDV) evolutiivisesta siirtoreitistä. Katso viitteet tekstistä.
Lisäksi CDV:n maantieteellisen alkuperän ja divergoitumisajankohdan epistemologisesta ja/tai tieteellisestä merkityksestä on olemassa tärkeitä johtolankoja, jotka on selvitettävä, jotta voidaan paremmin ymmärtää CDV:iden nykyistä vaikutusta lajien väliseen siirtymiseen, eläinten suojeluun ja zoonoosipotentiaaliin (kuva 1). On selvää, että toisin kuin MeV-infektio, jota ylläpitää yksi ainoa isäntä (ihminen), CDV on laajalti osoittautunut lupaavaksi patogeeniksi, joka aiheuttaa infektioita/sairauksia laajalle joukolle lihansyöjä- ja muita kuin lihansyöjälajeja (Martinez-Gutierrez ja Ruiz-Saenz, 2016). Tämä promiscuiteetti on johtunut paitsi CDV:n hemagglutiniinin (H) kyvystä olla vuorovaikutuksessa isäntäsolujen reseptorien kanssa, kuten SLAM mononukleaarisoluissa ja nektiini-4 epiteelisoluissa, myös edellä mainittujen reseptorien lajisekvenssien samankaltaisuudesta (Rendon-Marin ym., 2019). Nisäkkäiden SLAM-reseptorien, myös merinisäkkäiden, aminohappojen samankaltaisuus on >80 % (Ohishi ym., 2014), mikä tukee tuloksia lajien välisestä siirtymisestä. Lisäksi ihmisten, hiirien ja koirien välillä ei ole lajikohtaista vaihtelua nektiini-4-sekvensseissä, koska ihmisen nektiini-4 voisi toimia CDV:n in vitro -reseptorina (Noyce ym., 2011).
Luonnolliset CDV-taudinpurkaukset erilaisissa kädellisissä eläimissä ovat herättäneet huolta CDV:n mahdollisesta tarttumisesta ihmisiin (Yoshikawa ym., 1989; Sun ym., 2010; Qiu ym., 2011; Sakai ym., 2013a). On raportoitu, että CDV-apinakannoilla on luontainen kyky käyttää ihmisen nektiini-4:ää viruksen sisäänpääsyyn ja että nämä apinoiden CDV:t mukautuvat helposti käyttämään ihmisen CD150 (SLAM) -reseptoria viruksen H-proteiiniin tehtyjen minimaalisten aminohappomuutosten jälkeen (Bieringer ym., 2013; Sakai ym., 2013b). Cynomolgus-makakoiden (Macaca fascicularis) in vivo kokeellisen CDV-infektion perusteella MeV-immuniteetin läsnä ollessa makakit olivat kuitenkin osittain ristikkäissuojattuja CDV-haasteelta (De Vries et al., 2014). Tämä viittaa siihen, että vaikka CDV voi helposti tartuttaa kädellisiä, MeV-immuniteetti suojaa ja että CDV-infektio voi olla itsestään rajoittuva. Kun tämä tulos siirretään ihmisiin, on olemassa potentiaalinen CDV-infektion riski ihmisillä, joilla ei ole ristiin suojaavaa MeV-immuniteettia rokottamatta jättämisen ja rokotteiden epäonnistumisen vuoksi (Haralambieva ym., 2015) tai rokotusten puuttumisen vuoksi mahdollisella hävittämisen jälkeisellä aikakaudella (Holzmann ym., 2016).
”Nousevien virusten” on raportoitu voivan syntyä virusten siirtyessä eläimistä ihmisiin lajien välisenä siirtymisenä toisista lajeista toiseen (Wolfe ym., 2007). Uudet tutkimukset, sekä rakenteelliset että bioinformatiikan tutkimukset, viittaavat siihen, että vain yksi ainoa aminohappomuutos proteiinisekvenssissä voisi riittää voittamaan rajoituksen solureseptorien käytössä kahden eri isännän, kuten ihmisen ja märehtijän, välillä (Abdullah et al., 2018). Ainutlaatuinen mutaatio CDV:n H-proteiinissa in vitro mahdollistaa sen, että tämä patogeeni voi infektoida soluja, jotka ilmentävät ihmisen SLAM-reseptoria (Otsuki ym., 2013). Lisäksi, jos hyväksymme hypoteesin, jonka mukaan CDV kehittyi MeV:stä, voisi olla mahdollista, että CDV:n jälkeläinen voisi pystyä infektoimaan ihmisiä uudelleen, koska sekä viruksen että ihmisen evoluutio on jatkuvaa, kuten aiemmin on ehdotettu muissa malleissa, vaikka esi-isä ”hyppääjävirus” oli kadonnut maapallolta aikaa sitten (Emerman ja Malik, 2010).
Yksi mielenkiintoisimmista tuloksista, jonka Uhl ym. esittivät, on lisäksi sekä CDV:n että MeV:n geenien optimointi ihmisen codon usage biasiin (CUB), mikä viittaa siihen, että CDV:n codonien käyttö on lähempänä ihmisen CUB:ia kuin koiran CUB:ia, koska virus tai sen kantaisä, todennäköisimmin MeV, sopeutui alunperin ihmiseen (Uhl et al., 2019). CUB viittaa ilmiöön, jossa joitakin synonyymejä koodoneja käytetään useammin kuin toisia ja siihen, miten tämä preferenssi vaihtelee lajien sisällä ja lajien välillä (Behura ja Severson, 2013). RNA-viruksissa kodonien käyttö on valikoitumisen kohteena, koska virukset ovat täysin riippuvaisia isännän tRNA:sta ja bias johtuu siitä, että virukset vastaavat isäntänsä kodonien käyttöä (Jenkins ja Holmes, 2003). Evoluutio voi joskus suosia viruksia, jotka vastaavat isäntänsä kodoninkäyttöä edistääkseen replikaationopeutta ja sopeutumista isäntään, kuten on raportoitu muissa RNA-viruksissa (Goni ym., 2012; Lauring ym., 2012; Di Paola ym., 2018; Freire ym., 2018).
Viimeiseksi haluaisimme väittää, että joitain muitakin tekijöitä on otettava huomioon CDV:n mahdollisessa zoonoottisessa skenaariossa. MeV:n ja CDV:n välinen ristineutralisaatio on tunnustettu jo useiden vuosien ajan (Brown ja Mccarthy, 1974), ja tämä lähtökohta on ollut olemassa jo yli puolen vuosisadan ajan, kun MeV-rokotetta käytettiin suojaamaan pentuja CDV:tä vastaan iässä, jolloin emän passiivinen immuniteetti usein häiritsi CDV-rokotusta (Baker ym., 1966; Brown ym., 1972). Kaupallisen kaksois-CDV/MeV-rokotteen käyttöä suositellaan kuitenkin edelleen rokottamiseen äidin immuniteetin vallitessa, ja rokote on ollut hyödyllinen kliinistä tuhkarokkotautia vastaan kädellisillä (Christe ym., 2019). Näin ollen voidaan spekuloida, että MeV-laumaimmuniteetti estää CDV:n hyppäämisen ja mahdollisen uudelleen sopeutumisen ihmisiin koirien tai luonnonvaraisten eläinten välityksellä tapahtuvan tartunnan kautta.
Loppuhuomautukset
Viruspatogeenien evoluutiota ja alkuperää ei voida tutkia helposti; tämän jälkeen tarvitaan monitieteistä lähestymistapaa, jotta voidaan ymmärtää ja ehkä ennustaa uusia mahdollisia ihmisiin kohdistuvia virusuhkia. Erikoisen biologiansa vuoksi CDV:n kaltaiset viruspatogeenit edustavat ainutlaatuista mallia, jonka avulla voidaan ymmärtää lajien välisiä hyppyjä ja virusaiheuttajien zoonoosipotentiaalia hyvin lähellä ihmisväestöä. Perinteisten molekyylifylogeneettisten tutkimusten ja paleopatologisten töiden lisäksi tutkijoiden on omaksuttava erilaisia lähestymistapoja tutkiakseen CDV:n alkuperää ja nykyisiä virus- ja isäntävaatimuksia lajien väliselle hyppäämiselle. Laskennallisten menetelmien, kuten rakenteellisen bioinformatiikan ja paleovirologisten tutkimusten, käyttöönotto voisi auttaa ennustamisessa ja ennaltaehkäisyssä tai ainakin tarjota paremman ymmärryksen tästä kehittyvästä ja ehkä zoonoottisesta taudista erilaisesta näkökulmasta, jossa otetaan huomioon sekvensointidatan lisäksi myös rakenteet ja toiminnot keskeisinä tietoina tämän tavoitteen saavuttamiseksi.
Tekijöiden panos
Kaikki luettelossa mainitut kirjailijat ovat osallistuneet merkittävällä tavalla suoraan ja henkisesti työhön, ja he ovat myös hyväksyneet teoksen julkaistavaksi.
Rahoitus
Tätä työtä on taloudellisesti tukenut Departamento Administrativo de Ciencia, Tecnología e Innovación-COLCIENCIASin apuraha nro 123171249669 JR-S:lle.
Conflict of Interest Statement
Tekijät ilmoittavat, että tutkimus suoritettiin ilman kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdolliseksi eturistiriidaksi.
Brown, A. L., ja Mccarthy, R. E. (1974). Tuhkarokko- ja penikkatautivirusten välinen suhde määritettynä koirien viivästyneen tyypin yliherkkyysreaktioilla. Nature 248, 344-345. doi: 10.1038/248344a0
PubMed Abstract | CrossRef Full Text | Google Scholar