Morbilliviruses are highly contagious pathogens and are responsible for various outbreaks in unexposed populations (Pfeffermann et al., 2018). Acestea aparțin ordinului Mononegavirales și familiei Paramyxoviridae și sunt caracterizate de un genom ARN nesegmentat, liniar, cu catenă negativă (Lamb și Parks, 2013). Morbillivirușii se disting prin faptul că provoacă boli respiratorii, gastrointestinale, imunosupresoare și/sau neurologice de intensitate moderată până la severă la o gamă largă de gazde, inclusiv la oameni (virusul rujeolei), la carnivore (morbillivirusul canin, anterior virusul bolii de Carré), la bovine (virusul pestei bovine), la delfini și marsuini, precum și la alte specii de animale sălbatice pe cale de dispariție (Lamb și Parks, 2013; Martinez-Gutierrez și Ruiz-Saenz, 2016).
Virusul Measles (MeV) și morbillivirusul canin (CDV) sunt considerate cele mai contagioase virusuri din această familie (De Vries et al., 2015), iar din cauza potențialului ridicat de transmitere a CDV, precum și a potențialului său de transmitere între specii, autoritățile mondiale din domeniul sănătății, precum și cele din domeniul conservării sunt foarte preocupate de rolul CDV asupra conservării speciilor pe cale de dispariție și de posibilul „salt” de la animale la oameni (Terio și Craft, 2013; Ohishi et al., 2014). Câinii domestici sunt principala gazdă pentru CDV și ar putea fi considerați, de asemenea, un rezervor pentru alte mamifere (Suzuki et al., 2015; Duque-valencia et al., 2019); cu toate acestea, pe baza biologiei CDV, oamenii s-ar putea transforma, de asemenea, într-o potențială țintă (Cosby și Weir, 2018; Rendon-Marin et al, 2019).
În încercarea de a înțelege riscul potențial de transmitere a CDV la om, este necesar să se adune toate dovezile existente; iar studiul originii și diseminării acestui agent în populația canină ar putea prezenta o cheie importantă pentru înțelegerea acestui proces. Recent, un articol publicat în International Journal of Paleopathology a invitat la o discuție privind originea evolutivă a CDV. Acesta concluzionează că CDV a luat naștere ca agent patogen pandemic în America de Sud în urma infecției și adaptării MeV la câini în timpul perioadei de colonizare a Americii de Sud. Acest rezultat a fost obținut printr-o abordare interdisciplinară adoptată prin sintetizarea unei analize paleopatologice a 96 de câini precolumbieni (750-1470 CE) din situl Weyanoke Old Town, Virginia, cu rapoarte istorice, analize moleculare și epidemiologie morbillivirală (Uhl et al, 2019).
În mod notabil, populațiile de câini nativi din America aproape au dispărut după perioada de colonizare, iar câinii europeni și eurasiatici au fost introduși pe continent, lăsând puțin fond genetic al predecesorilor săi americani (Ni Leathlobhair et al., 2018). Un alt factor important care merită luat în considerare este faptul că ar fi putut fi introduse și boli „necunoscute”, ceea ce face mai dificilă urmărirea originii noilor agenți patogeni. Mai mult, presiunea de selecție artificială asupra câinilor domestici și chiar a populațiilor umane, în special în timpul perioadei de colonizare, ar fi putut spori incidența bolilor, limitând astfel variația genetică (Ostrander et al, 2017), ceea ce, la rândul său, ar putea însemna un răspuns mai puțin eficient împotriva agenților patogeni.
Printre acești agenți patogeni/bolile „noi”, CDV a fost descris pentru prima dată de Antonio de Ulloa y de la Torre-Giral în 1746 ca o boală care afecta câinii din regiunea Quito și din alte părți ale Americii de Sud și a fost raportat la scurt timp după aceea în Europa. CDV a fost înregistrat în Spania în 1760, cu 900 de decese într-o singură zi la Madrid, iar 3 ani mai târziu, adică în 1764 și 1770, a ajuns în Marea Britanie și, respectiv, Italia (Blancou, 2004). Transmisibilitatea virusului și sensibilitatea mai mare a cățeilor în comparație cu cea a câinilor adulți au fost raportate ulterior de Edward Jenner la începutul anilor 1800. El a comparat transmisibilitatea lor cu cea a MeV și a descoperit că supraviețuitorii erau protejați de infecții ulterioare (Jenner, 1809; Nambulli et al., 2016).
Privind pe scurt, după sosirea pionierilor europeni în secolul al XV-lea, noile boli infecțioase au devenit, fără îndoială, cea mai devastatoare consecință a colonizării, deoarece populațiile indigene americane nu au avut o expunere anterioară la agenții patogeni care deveniseră comuni în Europa (Walker et al., 2015). Prin urmare, multiplele epidemii de rujeolă au devastat populațiile indigene americane (Walker et al., 2015; Nambulli et al., 2016). Uhl et al. prin intermediul unei abordări mixte a dovezilor paleopatologice, istorice, moleculare și epidemiologice, au raportat că epidemiile severe de MeV în populațiile indigene americane au facilitat saltul MeV la populațiile mari de câini domestici din mediile urbane din America de Sud și adaptarea virusului ca CDV endemic (Uhl et al., 2019). De asemenea, înregistrările istorice ar putea dovedi că, la câțiva ani după această adaptare la câinii din America de Sud, CDV a fost transportat în Europa în 1760, unde a indus inițial epidemii extinse cu mortalitate ridicată înainte de a deveni endemic (Jenner, 1809).
Cu toate acestea, filogeografia moleculară legată de predicțiile evolutive și timpul până la cel mai recent strămoș comun (tMRCA) au fost calculate pentru originea CDV în Statele Unite în anii 1880 (cea mai mare densitate posterioară de 95%, 1858-1913) (Panzera et al., 2015), ceea ce contrazice în mod clar descrierea virusului în Europa în secolul al XVIII-lea. Analizele secvențelor care au condus la această ipoteză trebuie să fie examinate cu atenție din cauza prejudecăților și a disponibilității limitate a secvențelor care au fost utilizate în această reconstrucție filogeografică moleculară. În plus, multe secvențe ancestrale originale au fost pierdute din cauza labilității genomului ARN viral al CDV și al altor morbillivirusuri. Acești factori au dat naștere la punerea sub semnul întrebării a utilității calculelor actuale ale tMRCA pentru virusurile ARN (Sharp și Simmonds, 2011; Nambulli et al., 2016).
Potrivit lui Uhl et al., morbillivirusul ar fi putut proveni de la bovine în jurul anului 376 î.Hr. în „vechiul continent” (Figura 1), iar domesticirea animalelor ar fi putut avea o influență semnificativă asupra evenimentelor interspecii, urmărind probabil un punct de plecare în apariția MeV la aproximativ 900 AC (Uhl et al., 2019). Spre deosebire de reconstrucțiile filogenetice actuale ale CDV, divergența MeV este puternic susținută de analiza filogenetică bayesiană cu ceas relaxat. S-a demonstrat că timpul de divergență dintre MeV și virusul pestei bovine a avut loc aproximativ în secolele al XI-lea – al XII-lea (Furuse et al., 2010). Alte date moleculare, cum ar fi prezența unui nou morbillivirus (strâns înrudit cu CDV și PDV) care circulă la liliecii din Brazilia (DrMV), permit speculația că CDV și DrMV ar putea avea un strămoș comun sud-american (Drexler et al., 2012), susținând astfel indirect ideea originii sud-americane timpurii a CDV.
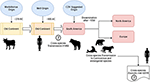
Figura 1. Reprezentarea schematică a posibilei căi de transmitere evolutivă a morbillivirusului canin (CDV). A se vedea textul pentru referințe.
Dincolo de semnificația epistemologică și/sau științifică a originii geografice și a datei de divergență a CDV, există indicii importante care trebuie clarificate pentru a înțelege mai bine impactul actual al CDV asupra transmiterii interspecifice, a conservării animalelor și a potențialului zoonotic (Figura 1). Este clar că, spre deosebire de infecția cu MeV, care este întreținută de o singură gazdă (oamenii), s-a dovedit pe scară largă că CDV este un agent patogen promiscuu care provoacă infecții/bolile la o gamă vastă de specii carnivore și non-carnivore (Martinez-Gutierrez și Ruiz-Saenz, 2016). Această promiscuitate a fost atribuită nu numai capacității hemaglutininei (H) a CDV de a interacționa cu receptorii celulari ai gazdei, cum ar fi SLAM în celulele mononucleare și nectin-4 în celulele epiteliale, ci și similitudinii dintre secvențele speciilor de receptori menționați mai sus (Rendon-Marin et al., 2019). Similaritatea aminoacizilor între receptorii SLAM ai mamiferelor, inclusiv mamiferele marine, este >80% (Ohishi et al., 2014), susținând astfel rezultatele transmiterii între specii. În plus, există o lipsă de variație legată de specie în ceea ce privește secvențele nectin-4 între oameni, șoareci și câini, deoarece nectin-4 uman ar putea funcționa ca un receptor in vitro pentru CDV (Noyce et al., 2011).
Focare naturală a CDV la diferite primate non-umane a stârnit îngrijorare cu privire la posibila transmitere a CDV la om (Yoshikawa et al., 1989; Sun et al., 2010; Qiu et al., 2011; Sakai et al., 2013a). Există rapoarte conform cărora tulpinile de maimuțe CDV au capacitatea intrinsecă de a utiliza nectin-4 uman pentru intrarea virusului și că aceste CDV-uri de maimuță se adaptează cu ușurință pentru a utiliza receptorul CD150 uman (SLAM) în urma unor modificări minime de aminoacizi ale proteinei H virale (Bieringer et al., 2013; Sakai et al., 2013b). Cu toate acestea, pe baza infecției experimentale in vivo cu CDV a macacilor Cynomolgus (Macaca fascicularis) în prezența imunității MeV, macacii au fost parțial protejați încrucișat de provocarea CDV (De Vries et al., 2014). Acest lucru sugerează că, deși CDV poate infecta cu ușurință primatele, imunitatea MeV este protectoare și că infecția cu CDV ar putea fi autolimitată. Transferând acest rezultat la oameni, există un risc potențial de infectare cu CDV la persoanele cărora le lipsește imunitatea MeV de protecție încrucișată din cauza nevaccinării și a eșecurilor vaccinale (Haralambieva et al., 2015) sau din cauza absenței vaccinării în posibila eră post-eradicare (Holzmann et al., 2016).
„Virușii emergenți” ar putea apărea, conform rapoartelor, prin intermediul transmiterii interspecifice a virusurilor de la animale la oameni (Wolfe et al., 2007). Studii noi, atât structurale, cât și bioinformatice, sugerează că o singură modificare a unui singur aminoacid într-o secvență proteică ar putea fi suficientă pentru a depăși restricția de utilizare a receptorilor celulari între două gazde diferite, cum ar fi oamenii și rumegătoarele (Abdullah et al., 2018). O mutație unică în proteina H a CDV in vitro permite acestui agent patogen să infecteze celulele care exprimă receptorul SLAM uman (Otsuki et al., 2013). Mai mult, dacă adoptăm ipoteza conform căreia CDV a evoluat din MeV, ar fi posibil ca un descendent al CDV să fie capabil să reinfecteze oamenii din cauza evoluției continue atât a virusului, cât și a oamenilor, așa cum s-a sugerat anterior în alte modele, chiar dacă „virusul săritor” ancestral a dispărut de mult timp de pe Pământ (Emerman și Malik, 2010).
În plus, unul dintre cele mai interesante rezultate prezentate de Uhl et al. este optimizarea atât a genelor CDV, cât și a MeV în funcție de prejudecata utilizării codonilor (CUB) la om, sugerând că utilizarea codonilor de către CDV este mai aproape de CUB uman decât de CUB canin, deoarece virusul sau progenitorul său, cel mai probabil MeV, a fost inițial adaptat la om (Uhl et al., 2019). CUB se referă la fenomenul în care unii codoni sinonimi sunt utilizați mai des decât alții și la modul în care această preferință variază în cadrul și între specii (Behura și Severson, 2013). În cazul virusurilor ARN, utilizarea codonilor este supusă selecției, deoarece virusurile sunt complet dependente de ARNt al gazdei, iar prejudecata rezultă din faptul că virusurile se potrivesc cu utilizarea codonilor gazdelor lor (Jenkins și Holmes, 2003). Evoluția poate favoriza uneori virușii care se potrivesc cu utilizarea codonilor gazdei lor pentru a promova viteza de replicare și adaptarea la gazdă, așa cum s-a raportat în cazul altor virusuri ARN (Goni et al., 2012; Lauring et al., 2012; Di Paola et al., 2018; Freire et al., 2018).
În cele din urmă, am dori să susținem că trebuie luați în considerare și alți factori în posibilul scenariu zoonotic al CDV. Neutralizarea încrucișată între MeV și CDV a fost recunoscută de mulți ani (Brown și Mccarthy, 1974), iar această premisă există de mai bine de o jumătate de secol, când vaccinul MeV a fost utilizat pentru a proteja puii împotriva CDV la o vârstă la care imunitatea maternă pasivă a interferat adesea cu vaccinarea împotriva CDV (Baker et al., 1966; Brown et al., 1972). Cu toate acestea, utilizarea unui vaccin comercial dublu CDV/MeV este încă recomandată pentru vaccinarea în prezența imunității materne, iar vaccinul a fost util împotriva bolii clinice de rujeolă la primatele neumane (Christe et al., 2019). Prin urmare, se poate specula că imunitatea de turmă MeV evită saltul CDV și posibila readaptare la om prin transmiterea prin intermediul câinilor sau al animalelor sălbatice.
Observații finale
Evoluția și originea agenților patogeni virali nu pot fi studiate cu ușurință; în continuare, este necesară o abordare multidisciplinară pentru a înțelege și, poate, pentru a prezice noi posibile amenințări virale pentru oameni. Datorită biologiei lor specifice, agenții patogeni virali, cum ar fi CDV, reprezintă un model unic pentru a înțelege saltul interspecii și potențialul zoonotic al agenților virali foarte apropiați de populația umană. Pe lângă studiile filogenetice moleculare tradiționale și lucrările de paleopatologie, cercetătorii trebuie să adopte abordări diferite pentru a studia originea CDV și cerințele actuale ale virusului și ale gazdei pentru saltul interspecii. Introducerea metodelor computaționale, cum ar fi bioinformatica structurală și studiile de paleovirologie, ar putea ajuta la predicția și prevenirea sau cel puțin ar putea oferi o mai bună înțelegere a acestei boli emergente și, probabil, zoonotice dintr-o perspectivă diferită, luând în considerare nu numai datele de secvențiere, ci și structurile și funcțiile ca informații cheie în acest scop.
Contribuții ale autorilor
Toți autorii enumerați au avut o contribuție substanțială, directă și intelectuală la această lucrare și au aprobat-o pentru publicare.
Finanțare
Această lucrare a fost susținută financiar de Departamento Administrativo de Ciencia, Tecnología e Innovación-COLCIENCIAS Grant nr. 123171249669 pentru JR-S.
Declarație privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Brown, A. L., și Mccarthy, R. E. (1974). Relația dintre virusul rujeolei și distemperul canin determinată de reacțiile de hipersensibilitate de tip întârziat la câini. Nature 248, 344-345. doi: 10.1038/248344a0
PubMed Abstract | Refef Full Text | Google Scholar
.