DNA
DNA je polymer, který se skládá z monomerů zvaných deoxynukleotidy. Monomer obsahuje jednoduchý cukr (deoxyribózu, zobrazenou níže černě), fosfátovou skupinu (červeně) a cyklickou organickou skupinu R (modře), která je analogická postrannímu řetězci aminokyseliny.
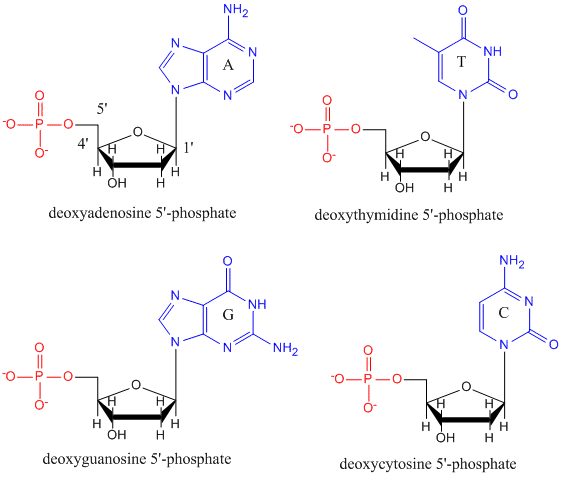
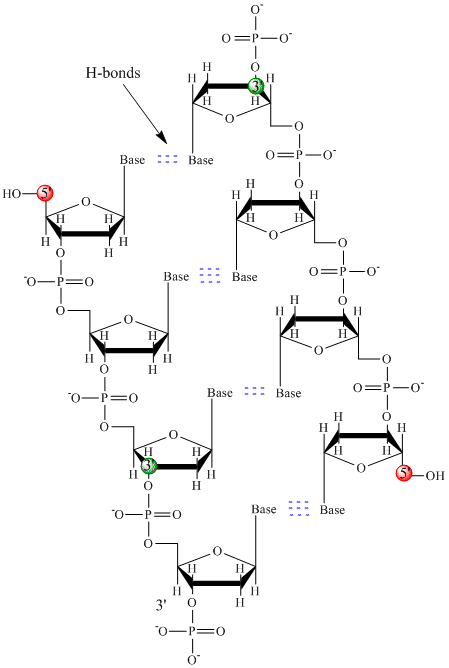
V DNA se používají pouze čtyři báze (na rozdíl od 20 různých postranních řetězců v bílkovinách), které budeme pro zjednodušení zkracovat na A, G, C a T. Jsou to báze, protože obsahují aminoskupiny, které mohou přijímat protony. Polymer se skládá z páteře cukr – fosfát – cukr – fosfát, přičemž ke každé molekule cukru je připojena jedna báze. Stejně jako u bílkovin je páteř DNA polární, ale také nabitá. Jedná se o polyaniont. Báze, analogické postranním řetězcům aminokyselin, jsou převážně polární. Vzhledem k nabité povaze páteře lze očekávat, že se DNA nesloží do kompaktního kulovitého (sférického) tvaru, i když se na polymer vážou kladně nabité kationty, jako je Mg, a stabilizují náboj. Místo toho DNA obvykle existuje jako dvouřetězcová (ds) struktura s cukr-fosfátovými páteřemi dvou různých řetězců probíhajících v opačných směrech (5′-3′ a druhý 3′-5′). Vlákna drží pohromadě vodíkové vazby mezi bázemi na komplementárních vláknech. DNA má tedy stejně jako bílkoviny sekundární strukturu, ale v tomto případě nejsou vodíkové vazby uvnitř páteře, ale mezi „postranními řetězci“ bází na protilehlých vláknech. Ve skutečnosti je nesprávné nazývat dsDNA molekulou, protože se ve skutečnosti skládá ze dvou různých komplementárních vláken držených pohromadě vodíkovými vazbami. Struktura ds-DNA znázorňující opačnou polaritu vláken je znázorněna níže.
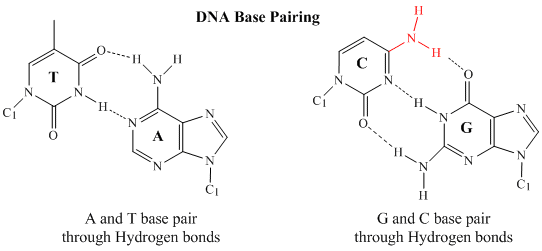
V dvouvláknové DNA může báze guanin (G) na jednom vlákně vytvořit tři H-vazby s bází cytosin (C) na druhém vlákně (nazývá se GC bázový pár). Báze thyminu (T) na jednom vlákně může vytvořit dvě H-vazby s bází adeninu (A) na druhém vlákně (tento pár se nazývá AT báze). Dvouřetězcová DNA má pravidelnou geometrickou strukturu s pevnou vzdáleností mezi oběma páteřemi. To vyžaduje, aby se páry bází skládaly z jedné báze s dvoukroužkovou (bicyklickou) strukturou (tyto báze se nazývají puriny) a jedné s jednokroužkovou strukturou (tyto báze se nazývají pyrimidiny). Proto G a A nebo T a C nejsou možnými partnery páru bází.
Dvojvláknová DNA se liší délkou (počtem spojených cukr-fosfátových jednotek), složením bází (kolik je každé sady bází) a sekvencí (pořadím bází v páteři). Následující odkazy poskytují interaktivní modely dsDNA v Jmol, které vytvořili Angel Herráez, Univ. de Alcalá (Španělsko) a Eric Martz.
- Jmol model dsDNA s páry bází a H-vazbami
- Jmol model vláken DNA a šroubovicové páteře
- Jmol model konců DNA a rovnoběžek
Chromozomy se skládají z jedné dsDNA s mnoha různými vázanými proteiny. Lidský genom má přibližně 3 miliardy párů bází DNA. Každý jednotlivý chromozom z páru má tedy v průměru asi 150 milionů párů bází a spoustu navázaných proteinů. dsDNA je vysoce nabitá molekula a lze na ni v prvním přiblížení pohlížet jako na dlouhou tyčinkovitou molekulu s velkým záporným nábojem. Jedná se o polyaniont. Tato velmi velká molekula musí být nějakým způsobem zabalena do malého jádra malé buňky. Ve složitých (eukaryotických) buňkách se tento problém balení řeší navíjením DNA kolem jádra komplexu čtyř různých párů (celkem osmi proteinů) histonových proteinů (H2A, H2B, H3 a H4), které mají čistý kladný náboj. Komplex histonového jádra s dsDNA navinutou přibližně 2,5krát se nazývá nukleosom.
Jmol model nukleosomu
DNA může nabývat dvou dalších typů dvoušroubovicových forem. Ta, kterou objevili Watson a Crick a kterou najdete ve většině učebnic, se nazývá B-DNA. V závislosti na aktuální sekvenci DNA a stavu hydratace DNA ji lze přimět k vytvoření dalších dvou typů dvoušroubovic, Z a A DNA. Forma A je mnohem otevřenější než forma B.
3,2 miliardy párů bází DNA u člověka obsahuje asi 24 000 krátkých úseků (genů), které kódují různé bílkoviny. Tyto geny jsou proloženy DNA, která pomáhá určit, zda je gen dekódován do molekul RNA (viz níže) a nakonec do proteinů. Aby se určitý gen aktivoval (neboli „zapnul“), musí se na oblast konkrétního genu navázat specifické bílkoviny. Jak mohou vazebné proteiny najít specifické vazebné cíle mezi obrovským množstvím párů bází, které mají v prvním přiblížení opakující se opakování cukr-fosfát-báze? Jmol níže ukazuje, jak lze dosáhnout specifičnosti. Když se DNA stáčí do dvojité šroubovice prostřednictvím párování bází mezi AT a GC, donory vodíkových vazeb (amidové Hs) a akceptory (Os) na bázích, které se nepoužívají při párování bází uvnitř řetězce,jsou stále k dispozici v hlavním a vedlejším háji šroubovice ds-DNA (viz Jmol níže). Unikátní sekvence párů bází budou vykazovat unikátní vzorce donorů a akceptorů vazeb H v hlavním háji. Tyto donory/akceptory mohou být rozpoznány specifickými vazebnými proteiny DNA, které po navázání mohou vést k aktivaci genu.
- Jmol model dsDNA zobrazující jedinečné donory a akceptory vazeb H v hlavním háji
.