ADN
ADN-ul este un polimer, format din monomeri numiți deoxinucleotide. Monomerul conține un zahăr simplu (deoxiriboză, reprezentată în negru mai jos), o grupare fosfat (în roșu) și o grupare organică ciclică R (în albastru) care este analogă cu lanțul lateral al unui aminoacid.
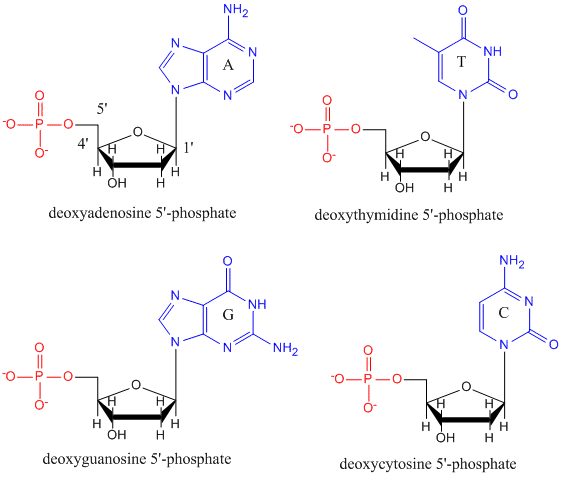
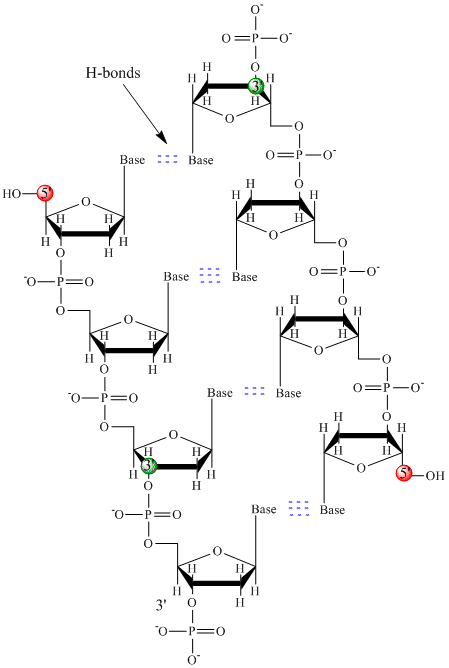
În ADN sunt folosite doar patru baze (spre deosebire de cele 20 de lanțuri laterale diferite din proteine) pe care le vom abrevia, pentru simplitate, ca A, G, C și T. Acestea sunt baze deoarece conțin grupe amină care pot accepta protoni. Polimerul este format dintr-o coloană vertebrală zahăr – fosfat – zahăr – fosfat, cu câte o bază atașată la fiecare moleculă de zahăr. Ca și în cazul proteinelor, coloana vertebrală a ADN-ului este polară, dar și încărcată. Este un polianion. Bazele, analoage lanțurilor laterale ale aminoacizilor, sunt predominant polare. Având în vedere natura încărcată a coloanei vertebrale, v-ați putea aștepta ca ADN-ul să nu se plieze într-o formă globulară (sferică) compactă, chiar dacă cationii încărcați pozitiv, cum ar fi Mg, se leagă de polimer și stabilizează sarcina acestuia. În schimb, ADN-ul există, de obicei, sub forma unei structuri bicatenare (ds), cu coloanele vertebrale de zaharuri-fosfați ale celor două șiruri diferite care merg în direcții opuse (5′-3′ și cealaltă 3′-5′). Șirurile sunt ținute împreună prin legături de hidrogen între bazele de pe șirurile complementare. Prin urmare, la fel ca și proteinele, ADN-ul are o structură secundară, dar în acest caz, legăturile de hidrogen nu se află în interiorul coloanei vertebrale, ci între bazele „lanțului lateral” de pe șirurile opuse. De fapt, este o denumire greșită să numim dsADN o moleculă, deoarece aceasta este formată în realitate din două șiruri diferite, complementare, ținute împreună prin legături de hidrogen. O structură de ADN-DS care arată polaritatea opusă a catenei este prezentată mai jos.
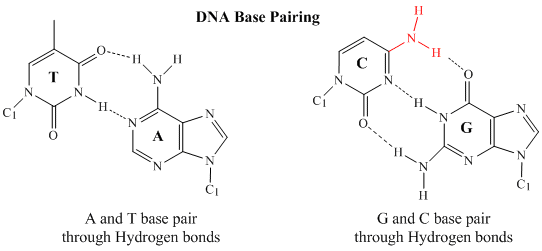
În ADN-ul dublu catenar, baza guanină (G) de pe o catenă poate forma trei legături H cu o bază citosină (C) de pe o altă catenă (aceasta se numește o pereche de baze GC). Baza timină (T) de pe un catenar poate forma două legături H cu o bază adenină (A) de pe celălalt catenar (aceasta se numește o pereche de baze AT). ADN-ul bicatenar are o structură geometrică regulată, cu o distanță fixă între cele două coloane vertebrale. Acest lucru impune ca perechile de baze să fie alcătuite dintr-o bază cu o structură cu două inele (biciclică) (aceste baze se numesc purine) și una cu o structură cu un singur inel (aceste baze se numesc pirimidine). Prin urmare, un G și un A sau un T și un C nu pot fi parteneri de perechi de baze.
ADN bicatenar variază în ceea ce privește lungimea (numărul de unități de zahăr-fosfat conectate), compoziția bazelor (câte din fiecare set de baze) și secvența (ordinea bazelor în coloana vertebrală). Următoarele linkuri oferă modele interactive Jmol de ADNDs realizate de Angel Herráez, Univ. de Alcalá (Spania) și Eric Martz.
- Model Jmol de ADN-DS cu perechi de baze și legături H
- Model Jmol de șiruri de ADN și coloana vertebrală elicoidală
- Model Jmol de capete de ADN și paralelisme
Cromozomii constau dintr-un ADN-DS cu multe proteine diferite legate. Genomul uman are aproximativ 3 miliarde de perechi de baze de ADN. Prin urmare, în medie, fiecare cromozom al unei perechi are aproximativ 150 de milioane de perechi de baze și o mulțime de proteine legate de el. dsADN este o moleculă foarte încărcată și poate fi privită, la o primă aproximație, ca o moleculă lungă, asemănătoare unei tije, cu o sarcină negativă mare. Este un polianion. Această moleculă foarte mare trebuie să fie cumva împachetată într-un mic nucleu al unei celule minuscule. În celulele complexe (eucariote), această problemă de împachetare este rezolvată prin înfășurarea ADN-ului în jurul unui complex de bază format din patru perechi diferite (opt proteine în total) de proteine histone (H2A, H2B, H3 și H4) care au sarcini pozitive nete. Complexul nucleu de histone cu ADN ds înfășurat de aproximativ 2,5 ori se numește nucleosom.
Modelul Jmol al nucleosomului
ADN-ul poate adopta alte două tipuri de forme dublu-helicoidale. Cea descoperită de Watson și Crick și regăsită în majoritatea manualelor se numește B-ADN. În funcție de secvența reală a ADN-ului și de starea de hidratare a ADN-ului, acesta poate fi convins să formeze alte două tipuri de elice bicatenară, Z și A ADN. Forma A este mult mai deschisă decât forma B.
Cele 3,2 miliarde de perechi de baze de ADN la om conțin aproximativ 24.000 de secțiuni scurte (gene) care codifică diferite proteine. Aceste gene sunt intercalate între ADN care ajută la determinarea dacă gena este decodificată în molecule de ARN (a se vedea mai jos) și, în cele din urmă, în proteine. Pentru ca o anumită genă să fie activată (sau „pornită”), anumite proteine trebuie să se lege de regiunea unei anumite gene. Cum pot proteinele de legare să găsească ținte de legare specifice printre numărul mare de perechi de baze care, la o primă aproximație, au o repetiție repetitivă zahăr-fosfat-bază? Jmol de mai jos arată cum poate fi obținută specificitatea. Atunci când ADN-ul se înfășoară într-o dublă helixă prin intermediul perechilor de baze între AT și GC, donatorii de legături de hidrogen (amida Hs) și acceptorii (Os) de pe bazele care nu sunt utilizate în împerecherea de baze intrastrand,sunt încă disponibile în șanțul major și minor al helixului ADN-DS (a se vedea Jmol de mai jos). Secvențele unice de perechi de baze vor prezenta modele unice de donatori și acceptori de legături H în canelura majoră. Acești donatori/acceptori pot fi recunoscuți de proteine specifice de legare a ADN-ului care, la legare, pot duce la activarea genei.
- Model Jmol de dsADN care prezintă donatori și acceptori unici de legături H în șanțul major