DNS
A DNS egy polimer, amely monomerekből, dezoxinukleotidokból áll. A monomer tartalmaz egy egyszerű cukrot (dezoxiribóz, lent fekete színnel ábrázolva), egy foszfátcsoportot (piros színnel) és egy ciklikus szerves R-csoportot (kék színnel), amely analóg az aminosavak oldalláncával.
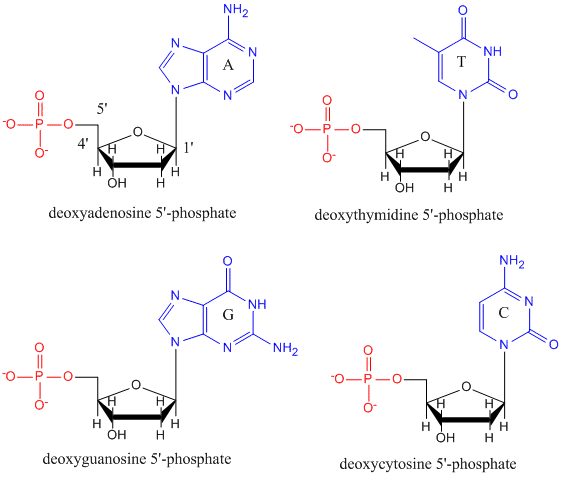
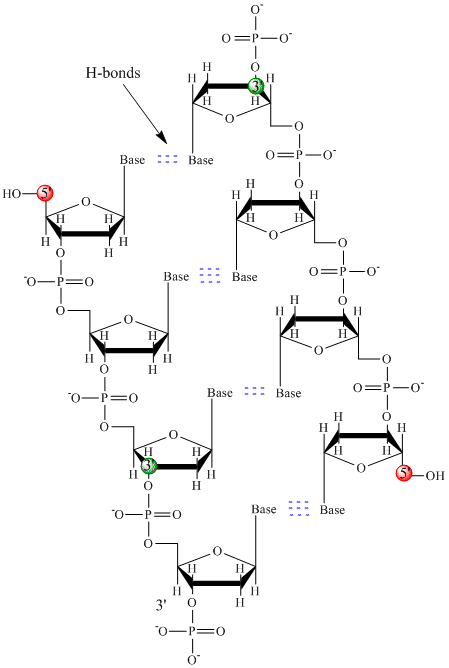
A DNS-ben csak négy bázis található (szemben a fehérjék 20 különböző oldalláncával), amelyeket az egyszerűség kedvéért A, G, C és T rövidítéssel jelölünk. Ezek bázisok, mivel protonok befogadására képes amincsoportokat tartalmaznak. A polimer egy cukor – foszfát – cukor – foszfát gerincből áll, ahol minden cukormolekulához egy-egy bázis kapcsolódik. A fehérjékhez hasonlóan a DNS gerincoszlop is poláris, de töltött. Ez egy polianion. A bázisok, az aminosavak oldalláncaihoz hasonlóan, túlnyomórészt polárisak. Tekintettel a gerincoszlop töltött jellegére, várható, hogy a DNS nem hajtódik kompakt gömb alakúvá (gömb alakúvá), még akkor sem, ha a pozitív töltésű kationok, mint például az Mg, kötődnek a polimer töltéséhez és stabilizálják a töltést. Ehelyett a DNS általában kettős szálú (ds) szerkezetben létezik, amelyben a két különböző szál cukor-foszfát gerincvonala ellentétes irányban fut (5′-3′ és a másik 3′-5′). A szálakat a komplementer szálak bázisai közötti hidrogénkötések tartják össze. A fehérjékhez hasonlóan tehát a DNS is rendelkezik másodlagos szerkezettel, de ebben az esetben a hidrogénkötések nem a gerincen belül, hanem az ellentétes szálak “oldalláncú” bázisai között vannak. A dsDNS-t valójában félrevezető molekulának nevezni, mivel valójában két különböző, komplementer szálból áll, amelyeket hidrogénkötések tartanak össze. A ds-DNS szerkezete, amely a szálak ellentétes polaritását mutatja, az alábbiakban látható.
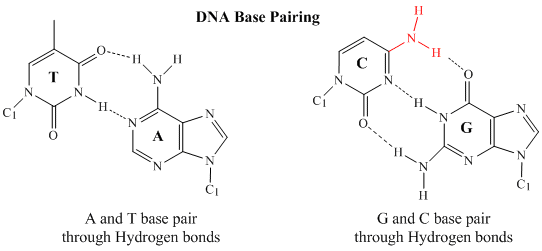
A kétszálú DNS-ben az egyik szálon lévő guanin (G) bázis három H-kötést tud kialakítani a másik szálon lévő citozin (C) bázissal (ezt nevezzük GC-bázispárnak). Az egyik szálon lévő timin (T) bázis két H-kötést tud kialakítani a másik szálon lévő adenin (A) bázissal (ezt AT bázispárnak nevezzük). A kettős szálú DNS szabályos geometriai szerkezettel rendelkezik, a két gerincoszlop között rögzített távolság van. Ez megköveteli, hogy a bázispárok egy kétgyűrűs (biciklikus) szerkezetű bázisból (ezeket a bázisokat purinoknak nevezzük) és egy egygyűrűs szerkezetű bázisból (ezeket a bázisokat pirimidineknek nevezzük) álljanak. Ezért a G és az A vagy a T és a C nem lehetséges bázispár partnerek.
A kétszálú DNS hossza (az összekapcsolt cukor-foszfát egységek száma), bázisösszetétele (az egyes báziscsoportokból mennyi van) és szekvenciája (a bázisok sorrendje a gerincoszlopban) változik. Az alábbi linkeken a dsDNS interaktív Jmol modelljei találhatók, amelyeket Angel Herráez, Univ. de Alcalá (Spanyolország) és Eric Martz készített.
- ds-DNS Jmol modellje bázispárokkal és H-kötésekkel
- Jmol modell a DNS szálakról és a spirális gerincről
- Jmol modell a DNS végekről és a párhuzamosokról
A kromoszómák egy dsDNS-ből állnak, amelyhez sok különböző fehérje kapcsolódik. Az emberi genom körülbelül 3 milliárd bázispár DNS-t tartalmaz. Ezért átlagosan egy kromoszómapár minden egyes kromoszómájához körülbelül 150 millió bázispár és rengeteg fehérje kötődik. dsDNS egy erősen töltött molekula, és első közelítésben egy hosszú, rúdszerű, nagy negatív töltéssel rendelkező molekulának tekinthetjük. Ez egy polianion. Ezt a nagyon nagy molekulát valahogy be kell pakolni egy aprócska sejtmagba. Az összetett (eukarióta) sejtekben ezt a pakolási problémát úgy oldják meg, hogy a DNS-t egy négy különböző pár (összesen nyolc fehérje) hisztonfehérje (H2A, H2B, H3 és H4) magkomplex köré tekerik, amelyek nettó pozitív töltéssel rendelkeznek. A körülbelül 2,5-szer körbetekert dsDNS-t tartalmazó hisztonmagkomplexet nukleoszómának nevezzük.
Jmol modell a nukleoszómáról
A DNS két másik típusú kettős hélix alakot is felvehet. A Watson és Crick által felfedezettet, amely a legtöbb tankönyvben megtalálható, B-DNS-nek nevezik. Az aktuális DNS-szekvenciától és a DNS hidratáltsági állapotától függően két másik típusú kettős hélixet, a Z és az A DNS-t is kialakíthatjuk. Az A forma sokkal nyitottabb, mint a B forma.
Az emberben található 3,2 milliárd bázispár DNS körülbelül 24 000 rövid szakasz (gén) tartalmaz, amelyek különböző fehérjéket kódolnak. Ezek a gének a DNS között vannak elszórtan, ami segít meghatározni, hogy a gén RNS-molekulákká (lásd alább) és végül fehérjékké dekódolódik. Ahhoz, hogy egy adott gén aktiválódjon (vagy “bekapcsolódjon”), specifikus fehérjéknek kell kötődniük az adott gén adott régiójához. Hogyan találják meg a kötőfehérjék a specifikus kötődési célpontokat a rengeteg bázispár között, amelyek első közelítésben ismétlődő cukor-foszfát-bázis ismétlődést tartalmaznak? Az alábbi Jmol mutatja, hogyan érhető el a specifitás. Amikor a DNS kettős spirálba tekeredik az AT és GC közötti bázispárok révén, a ds-DNS spirál nagy és kis barázdájában még mindig rendelkezésre állnak hidrogénkötés-donorok (amid Hs) és akceptorok (Os) a bázisokon, amelyeket nem használnak fel a szálon belüli bázispárosodásban (lásd az alábbi Jmol-t). Az egyedi bázispár-szekvenciák a H-kötés donorok és akceptorok egyedi mintázatát mutatják a major grove-ban. Ezeket a donorokat/akceptorokat specifikus DNS-kötő fehérjék felismerhetik, amelyek a kötés hatására génaktiválódáshoz vezethetnek.
- A dsDNS Jmol modellje, amely egyedi H-kötés donorokat és akceptorokat mutat a major grove-ban
.